Environmental epidemiology, defined as the study of the spatial or spatiotemporal distribution of disease in relation to possible environmental factors, constitutes an important tool for better understanding the dynamics of parasitic infections and the development of suitable control and prevention strategies (Diggle 1993). GIS is a valuable tool of environmental epidemiology, but has not been extensively used globally in the study of disease distribution and dynamics, although epidemiological data clearly have spatial components. Perhaps the earliest and most celebrated example is John Snow's detection of the Broad Street water pump as the source of the 1840s cholera outbreak in London. He demonstrated that the association between cholera deaths and contaminated water supplies showed a striking geographic distribution (Henk et al. 1991; Diggle 1993).
In Tunisia, GIS have never been applied to health problems, although there has been geographic recognition of households and manual cartography for improving insecticide coverage within the context of malaria control programs. This approach, although very simple, constituted a valuable tool for the success of malaria control. Unfortunately, this technique was neither improved upon nor replicated for other parasitic diseases such as leishmaniasis and hydatid cystic, or other health hazards heavily influenced by the environment. In fact, this technology could not be sustained manually for two reasons:
This report presents the preliminary application of an automated GIS for the investigation of the spatiotemporal dynamics of ZCL in relation to potential environmental risk factors, in a pilot focus in Central Tunisia. More precisely, this work aims to:
The constitution of a spatially referenced database was performed using a personal computer to introduce two types of information. Nonlocational or descriptive data refer to the features or attributes (variables in the survey form describing individuals, animals or habitats). Locational or spatial data consist of lines (segments), such as the limits of the area, roads, and so on, points (nodes) corresponding to the dwellings, and areas (polygons) colonized by rodents. Spatial data were digitized into independent coverages of the three types using a format A0 digitizer (Summergraphics) in the GIS department of the Institut Regional des Sciences de l'Information et de Télécommunications (IRSIT, Tunis).
The epidemiological geographic information system (EPI-GIS) was realized using the GIS software package ARC/INFO 3.4D Plus (ESRI, Redlands). Thematic maps were printed using a format A0 plotter and an inkjet colour printer.
The absence of an interface for linking the descriptive database management software (ORACLE PC version) to the GIS software (ARC/INFO) required the conversion of the ORACLE data tables into a DBF format compatible with INFO, the database manager of ARC/INFO. Thus, it was possible to join the DBF descriptive data tables to the features attribute tables (AAT: arc attribute table; PAT: point/polygon attribute table) using the command JOIN.
The EPI-GIS conceived and developed by the laboratory of Ecology and Epidemiology of Parasitic Diseases (LEEP) team permitted the mapping of spatial distribution of ZCL over 10 years and to relate this distribution to the potential vegetation cover, particularly to the presence of chenopods. The command INTERSECT was used to overlay biotope coverages with the dwellings.
Buffers around a focal point believed to be the source of the epidemic were generated with the command BUFFER. This focal point corresponds to a cemetery where P. obesus colonies were confirmed in 1983 by field observations (Ben Ismail et al. unpublished data). Distances between this focal point and dwellings with cases were calculated using the command NEAR.
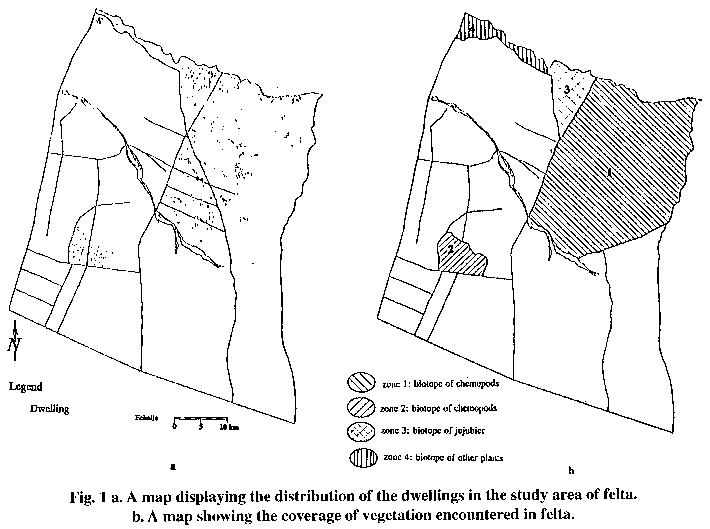
Overlay and intersection operations between different kinds of biotopes (I: presence of chenopods, II: presence of jujubier, III: presence of other plants) and clusters of dwellings located inside these biotopes were performed for the estimation of biotope-wise.
The distribution of the incidence of the infection shows two epidemic peaks in 1985 and 1987. The resulting graph shows neither the pattern of the spatial distribution of ZCL, nor its relation to possible environmental factors.
The distribution of the proportion of susceptibles Q(x) over the age groups (0-10 years) revealed a trend for decay. Assuming a constant exponential decay over the age groups (Henk et al. 1991), and using the log likelihood method, Q(x) was used to estimate the mean FOI for the whole period of time. It was found to be 0.017 per year, showing that the focus is hypoendemic for ZCL according to the Lysenko and Beljaev classification (1987).
Using the annual information, the number of immunes (new cases and previous cases) and the number of susceptibles, L(x) were estimated and the FOI was directly derived for the corresponding epidemiological year.
Table 1 shows the evolution of the force of infection through time and the different parameters derived from the database for its estimation. These results show that the FOI, far from being stable over time, follows the same trend as the incidence of infection with two peaks in 1985 and 1987.
| Table 1. Evolution of the force of infection through time and the parameters used for its estimation. | ||||
|---|---|---|---|---|
| Year | Susceptibles | ZCL Cases | Incidence L | Force of infection (lambda) |
| 1983 | 3334 | 15 | 0.004 | 0.005 |
| 1984 | 3444 | 64 | 0.018 | 0.019 |
| 1985 | 3489 | 392 | 1.112 | 0.119* |
| 1986 | 3258 | 53 | 0.099 | 0.016 |
| 1987 | 3351 | 332 | 0.099 | 0.104* |
| 1988 | 3142 | 69 | 0.021 | 0.022 |
| 1989 | 3183 | 73 | 0.022 | 0.023 |
| 1990 | 3215 | 21 | 0.006 | 0.007 |
| 1991 | 3315 | 37 | 0.011 | 0.011 |
| 1992 | 3486 | 24 | 0.006 | 0.007 |
| *Values of lambda corresponding to epidemic peaks in 1985 and 1987. | ||||
To visualize the spatial dynamics of the disease, an overlay of the base map of the area and the coverage of the dwellings without cases was achieved (Fig. 1a). The different biotopes encountered in the study area are displayed in Fig. 1b.
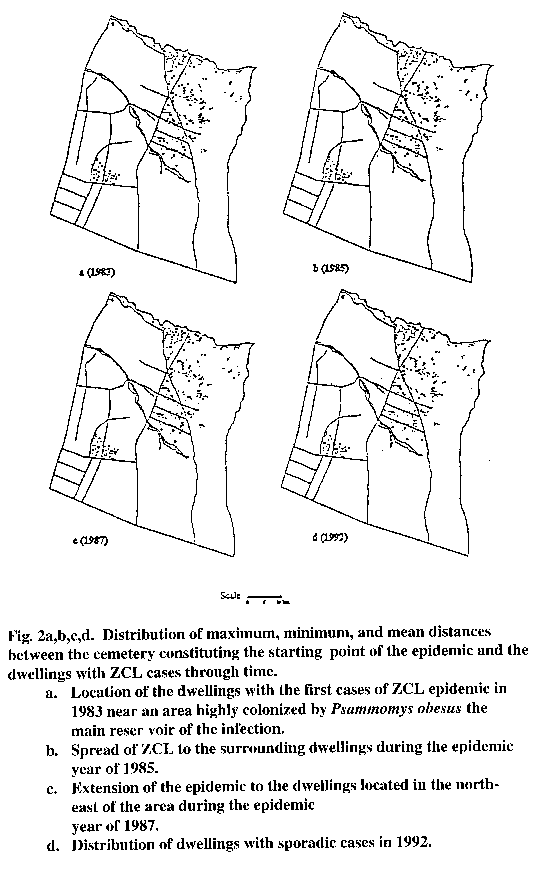
Figure 2a,b,c,d presents maps displaying the spatial distribution of dwellings with ZCL cases, for the years 1983, 1985, 1987, and 1992 respectively. The location of the first cases that occurred in 1983 (Fig. 2a), allowed the identification of the initial starting point of the epidemic in a cemetery that had been highly colonized by the P. obesus, the main reservoir of the parasite.
The capabilities of the EPI-GIS were applied to select clusters of dwellings according to the kind of biotope where they were located. This procedure permitted the comparison of the FOI in different biotopes during the epidemic years of 1985 and 1987 (Table 2). The results showed a large heterogeneity of the force of infection in the different selected biotopes, ranging from 0.23 in the biotope of chenopods to 0.01 in other biotopes in 1985, and confirmed that the distribution of ZCL is very sensitive to environmental factors.
| Table 2. Estimation of the force of infection (lambda) in different biotopes during the epidemic years (1985, 1987). | ||||||
|---|---|---|---|---|---|---|
| Biotope | Population size | Susceptibles | ZCL cases | Incidence | Force of infection (lambda) | Level of endemicity* |
| 1985 | ||||||
| 1 | 2578 | 2126 | 239 | 0.112 | 0.119 | méso |
| 2 | 373 | 340 | 71 | 0.209 | 0.234 | méso |
| 3 | 656 | 635 | 23 | 0.036 | 0.037 | hypo |
| 4 | 201 | 189 | 3 | 0.016 | 0.016 | hypo |
| 1987 | ||||||
| 1 | 2373 | 2309 | 354 | 0.153 | 0.166 | méso |
| 2 | 346 | 346 | 28 | 0.081 | 0.084 | méso |
| 3 | 606 | 597 | 1 | 0.002 | 0.002 | hypo |
| 4 | 185 | 185 | 9 | 0.049 | 0.05 | hypo |
| 1 and 2: biotope of chenopods with borrows of the rodent Psammomys obesus the main reservoir of ZCL. 3: biotope of jujubier with borrows of the rodent Meriones shawi secondary reservoir of ZCL. 4: other kinds of plantation having no relation with the reservoir of ZCL. * the level of endemicity is rated according to Lysenko and Beljaev classification. | ||||||
Indeed, in the biotopes 1 and 2 of chenopods the population of rodents is very dense, and individuals are exposed to a high risk of transmission. In these biotopes, the infection is mesoendemic, while it is hypoendemic in biotope 3, where the transmission is expected to be lower due to the absence of the infection reservoir. This analysis confirmed the heterogeneity of the transmission within the same small focus.
Buffers were generated every 200 m around the cemetery at the beginning of the epidemic in 1983, to estimate the evolution of the FOI around this starting point.
The distances between the starting point of the epidemic and the dwellings with cases ranged between 251 m and 1102 m in 1983. The maximum distance increased very quickly in 1984 to reach 2951 m, and remained almost unchanged over the following years.
The calculation of such classical epidemiological indicators usually includes the whole population surrounded by administrative boundaries of the district for deriving the indicator denominator (incidence, prevalence), assuming that all individuals are under an equal risk of infection. This assumption, although plausible for many health problems, is clearly misleading for vector-borne zoonosis where the abundance of both the vector and the reservoir is highly dependant upon the type of biotope where people are living. Within the context of ZCL, the EPI-GIS combined demographic, medical, and geographic data, and allowed the estimation of FOI through time and space. It also facilitated the study of relationships between the distribution and diffusion of the disease, and environmental risk factors. It offered the opportunity to test the plausibility of the basic assumptions of Lysenko and Beljaev's model (the homogeneity of the transmission through time and space) (1987). It was clearly demonstrated that these assumptions are unrealistic in the Tunisian context, raising the need for considering spatial heterogeneities and instability through time in the estimation of the force of infection.
GIS clearly confirmed the role of chenopods as determinants of transmission, and permitted the display of precise spatial distribution of infection. This result corroborates previous findings based on different epidemiological approaches (Ben Salah et al. 1994). GIS allowed the refinement of epidemiological parameters and the stratification of the studied area to different levels of risks for the transmission, which has very important implications in terms of understanding the determinants of the infection, its dynamics, and the allocation of resources for control.
By evaluating the distances between the source of the epidemic and the dwellings with cases, the reach of an epidemic of ZCL is estimated as approximately 1 km. Such information is extremely important for designing control measures based on environmental changes, such as ploughing the zones of chenopods. It informs precisely on the minimum area necessary to be ploughed to interrupt the transmission of the infection to people.
This approach appears to be more realistic than one based on the measurement of sandfly vector dispersal (maximum flight distance) (Killick-Kendrix et al. 1984). This research stimulated the collection of other environmental factors such as soil type, climatological parameters, waterways, and proximity to parasite sources to establish the relationships between these parameters and the spatial patterns of the disease.
In the context of ZCL, EPI-GIS provides a excellent framework within which the overlaying of a large number of geographic factors (coverages) and the association of them with the disease, the vector, and the reservoir can be undertaken. The GIS approach will become more important if the present research is extended to a larger geographic region. As more data on the trophic preferences and dynamic of the reservoir population become available, the EPI-GIS can assist epidemiologists to predict the spread and establishment of the disease in new locations which constitute a valuable tool for resource allocation and the implementation of preventive measures.
 Back to Contents -
Back to Contents -
 To next section
To next section
This file was created 23 February 1996
{kind=link}
{kind=link}